Guizhou Engineering Research Center of Industrial Key-technology for Dendrobium Nobile, Guizhou Engineering Research Center for Orchid Medicinal Plant Breeding and Efficient Application, Zunyi Medical University, Zunyi 563000, China
b.
State Key Laboratory of Discovery and Utilization of Functional Components in Traditional Chinese Medicine, Institute of Interdisciplinary Integrative Medicine Research, Shanghai University of Traditional Chinese Medicine, Shanghai 201203, China
c.
State Key Laboratory of Drug Research, Shanghai Institute of Materia Medica, Chinese Academy of Sciences, Shanghai 201203, China
Received Date:
26 February 2025 Accepted Date:
14 May 2025 Revised Date:
28 April 2025 Available Online:
15 April 2026
Abstract:
Bile salt hydrolase (BSH), a gatekeeper enzyme in bile acid metabolism, regulates the host's bile acid profile and is closely associated with various metabolic diseases. However, suitable methods for measuring its activity in living systems remain scarce. Herein, a novel far-red fluorogenic substrate (CA-ABEI) for BSH was designed and developed by conjugating cholic acid with an aminocoumarin fluorophore. Under physiological conditions, CA-ABEI can be rapidly hydrolyzed by BSH from various bacterial sources to form ABEI, triggering strong fluorescence enhancement at 620 nm. Specifically activated by BSH, CA-ABEI enables accurate detection of BSH activity in biospecimens, including pure enzymes, bacteria and intact fecal slurries, and the first bioimaging of BSH activity in both BSH-expressing engineered Escherichia coli and natural intestinal microbiota. Moreover, a high-throughput screening platform was established using CA-ABEI, enabling the evaluation of BSH inhibitory effects from 96 herbal extracts. Pu-erh tea emerged as a potent BSH inhibitor and its active components were subsequently characterized, aiding the discovery of novel BSH inhibitors. Collectively, CA-ABEI proved to be a powerful tool for monitoring BSH activity in complex biological systems with value for exploring physiological functions and rapid screening of inhibitors.
The gut microbiota, an extremely dynamic and chemically diverse community, plays a crucial role in nutrient acquisition, energy regulation, and the maintenance of human health, earning its recognition as an important "organ" [1]. Within this diverse microbial community, bile salt hydrolase (BSH)-producing bacteria have garnered significant interest for their pivotal role in regulating bile acid (BA) metabolic equilibrium [1–4]. BSH, also referred to as cholylglycine hydrolase, catalyzes the hydrolysis of amide bonds in conjugated BAs, releasing glycine or taurine and enabling the BA pool to undergo 7α-dehydroxylation, which generates secondary BAs [5,6]. Beyond their role in aiding the digestion and absorption of fat-soluble nutrients, BAs function as signaling molecules that interact with specific host receptors to modulate essential physiological functions [3,7]. For instance, BAs can influence a range of nuclear receptors and G protein-coupled receptors [8–10], such as the farnesoid X receptor (FXR), vitamin D receptor (VDR), pregnane X receptor (PXR), constitutive androstane receptor (CAR), and G protein-coupled bile acid receptor (GPBAR1 or TGR5). Dysregulation of these receptors can contribute to metabolic disturbances, potentially leading to conditions like hypercholesterolemia, inflammatory bowel disease (IBD), type 2 diabetes, gallstones, and colorectal cancer [11–16].
As a gatekeeper enzyme in BA metabolism [3,6], BSH regulates the first step of BA conversion in the intestine, thereby controlling the overall composition of the BA pool and ultimately influencing the gut microbiota composition. Recently, two articles published on the same day in Nature reported that BSH exhibits acyltransferase activity, generating a set of previously underappreciated bile acid metabolites with the potential to impact the microbiome and human health [17,18]. Extensive research has demonstrated that abnormal regulation of BSH activity is closely associated with various health conditions, such as metabolic dysfunctions, inflammatory responses, and infections. Therefore, developing detection methods for BSH activity in complex biological samples, as well as developing BSH regulators, is of great importance for both their therapeutic applications and the study of their biological functions in related diseases.
In recent years, fluorescent probes targeting bioactive proteins or enzymes in microorganisms have been developed, significantly enhancing the detection of enzymes and the discovery of bioactive molecules [19–21]. However, the availability of fluorogenic substrates for evaluating BSH activity in gut microbiota remains limited. This limitation arises from two main challenges: firstly, the complex chemical environment, heterogeneous distribution, and highly dynamic nature of the microbiota make it difficult to accurately measure BSH activity in the gastrointestinal tract. Secondly, bsh genes are dispersed across a wide range of bacterial species, many of which are unculturable with existing techniques. Notably, all existing BSH fluorogenic substrates have been developed based on BSH from a single bacterial strain, with no reports on substrates targeting BSH from multiple bacterial sources, such as CA-AMCA [22] and BAL [23] are targeting L. plantarum BSH, while ChoRhos is targeting L. plantarum BSH and L. gasseri BSH [24]. Furthermore, these substrates emit at short wavelengths (λem ≤ 525 nm), making them susceptible to biological matrix interference and reduced detection sensitivity (Table S1 in Supporting information).
To address these issues, we first expressed five recombinant BSHs from major bacterial sources [25]: Lactobacillus salivarius (Ls), Enterococcus faecalis (Ef), Bifidobacterium longum (Bl), Clostridium perfringens (Cp), and Bacteroides thetaiotaomicron (Bt) in E. coli BL21 (DE3) (Fig. S1 in Supporting information). Subsequently, building on the catalytic properties of BSH, a series of fluorogenic substrates were designed and synthesized, among which CA-ABEI exhibited the strongest fluorescence response across all five BSHs. CA-ABEI featured a conjugate of cholic acid (CA) and an aminocoumarin fluorophore, which is hydrolyzed by BSH to emit fluorescence at 620 nm upon 520 nm excitation (Fig. S2 in Supporting information). It exhibited excellent optical properties, remarkable stability, and exceptional sensitivity and specificity for BSH detection. CA-ABEI was successfully employed to accurately measure BSH activity in various biological samples, including purified enzymes, bacterial cultures, and intact fecal slurries. Furthermore, it facilitated high-throughput screening for BSH inhibitors and enabled the first bioimaging of BSH activity in both BSH-expressing engineered E. coli and natural intestinal microbiota. Given BSH's involvement in bacterial persistence and its implications for host health, CA-ABEI holds great promise as an effective tool for screening novel therapeutic agents and for advancing our understanding of BSH function in human health.
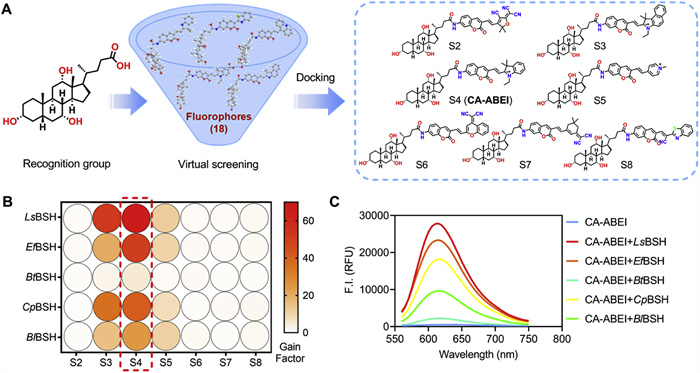
Coumarin-based fluorescent probes have emerged as preferred imaging tools to real-time and noninvasively monitor target analyte(s) in biological systems, due to their inherent merits of tunable optical properties, facile synthesis, excellent cell-membrane permeability, and favorable biocompatibility [26–30]. BSH is known to catalyze hydrolytic reactions of the C-24 amide bond of conjugated BAs into unconjugated BAs. Cholic acid has been confirmed as an effective recognized unit for constructing BSH substrates [22–24,31]. Based on the hydrolysis mechanism of BSH, eighteen coumarin-based far-red fluorogenic substrates were designed as BSH substrates by conjugating CA with diverse fluorophores (Fig. 1A and Fig. S3 in Supporting information). Considering the wide variety of BSH, crystal structures of five BSH enzymes (PDB ID: pdb:5HKE for LsBSH, 4WL3 for EfBSH, 6UFY for BtBSH, 2BJF for CpBSH and 2HF0 for BlBSH) were retrieved and docked with each fluorogenic substrate. Ensemble docking results indicated that S2–S8 exhibited favorable binding energies and short catalytic distances. Notably, for 5HKE and 2BJF, the binding energies were all below –12 kcal/mol, and the catalytic distances between the sulfhydryl group of Cys-2 (a key conserved amino acid in BSH) and the C-24 carbonyl carbon of each probe were all <5 Å (Table S2 in Supporting information). The results suggested that S2–S8 have more potential to serve as the substrates of BSH from multiple bacterial sources. Subsequently, these seven compounds were synthesized and fully characterized (Scheme S1 and Figs. S17–S56 in Supporting information), and their hydrolysis rates by BSH were assessed under physiological conditions. As shown in Fig. 1B, compounds S3–S5 were efficiently hydrolyzed, whereas S2 and S6–S8 were hardly hydrolyzed in the presence of the five BSHs. Among them, S4 demonstrated the highest gain factor towards five BSHs. The fluorescence emission spectrum exhibited a substantial intensification (67-fold) at 620 nm upon LsBSH treatment (Fig. 1C). Through structure-guided molecular design combined with virtual screening, S4 (CA-ABEI) was initially characterized as a universal far-red fluorogenic substrate for BSH.
Figure 1
Figure 1.
Rational design and screening of fluorogenic substrates for BSHs. (A) Design and virtual screening of BSH fluorogenic substrates. (B) The fluorescence response of S2–S8 (10 µmol/L) after reaction with five BSHs (20 µg/mL). (C) Fluorescence spectral changes of CA-ABEI (10 µmol/L) after reaction with five BSHs (20 µg/mL). λem = 520 nm, data are presented as mean ± SD (n = 3).
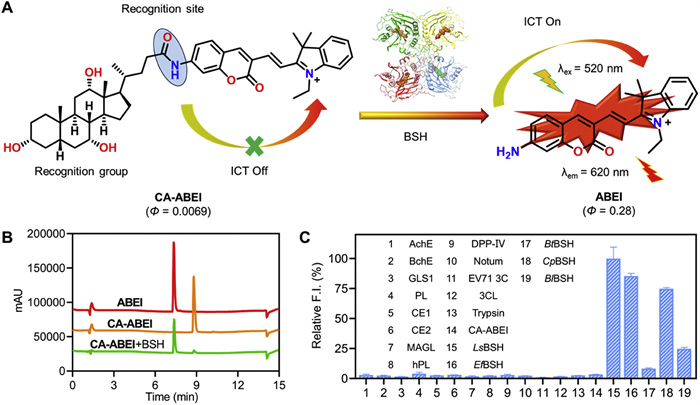
The spectral properties and sensing mechanism of CA-ABEI towards BSH were further investigated. Mechanistically, the amide linkage at the C-24 position of CA-ABEI served dual functions as both a fluorescence quencher and recognition element, effectively inhibiting the intramolecular charge transfer (ICT) mechanism within the coumarin-scaffold fluorophore. Upon BSH-mediated hydrolysis of the amide bond, the liberated amino group formed ABEI, a highly fluorescent product with a quantum yield of 0.28. This triggered an ICT process, resulting in strong fluorescence emission at 620 nm (Fig. 2A). The hydrolysis product ABEI was confirmed by matching its retention times on HPLC and the MS/MS spectra with those of the standards (Fig. 2B). Next, the specificity of CA-ABEI toward BSHs was evaluated by testing its reactivity with various hydrolases abundantly present in human tissues. As shown in Fig. 2C, CA-ABEI exhibited a rapid hydrolysis rate upon the addition of BSHs, while only negligible fluorescence signals were observed with other human hydrolases, indicating their lack of involvement in the hydrolysis of the CA-ABEI. Additionally, the potential interference of endogenous amino acids, metal ions and bioactive small molecules on the fluorescence detection of CA-ABEI was investigated. As demonstrated in Fig. S4A (Supporting information), none of the endogenous substances induced fluorescence enhancement of CA-ABEI. Although high concentrations of SO32⁻ and HSO3⁻ affected the fluorescence signal of ABEI, their influence is negligible under physiological conditions due to their low endogenous levels (Figs. S4B–D in Supporting information) [32]. Furthermore, ABEI displayed favorable photostability (70 min, Fig. S5 in Supporting information) and pH stability (pH 2–8, Fig. S6 in Supporting information). These findings demonstrated that CA-ABEI possesses exceptional specificity for BSHs over endogenous substances, making it a highly promising tool for monitoring BSH activity in biological systems.
Figure 2
Figure 2.
The sensing mechanism and enzyme specificity of CA-ABEI towards BSHs. (A) The sensing mechanism of CA-ABEI for BSH. (B) HPLC characterization of BSH—catalyzed hydrolysis of CA-ABEI. (C) The selectivity of CA-ABEI (10 µmol/L) among various hydrolases (10 µg/mL). λex = 520 nm, λem = 620 nm. Data are presented as mean ± SD (n = 3).
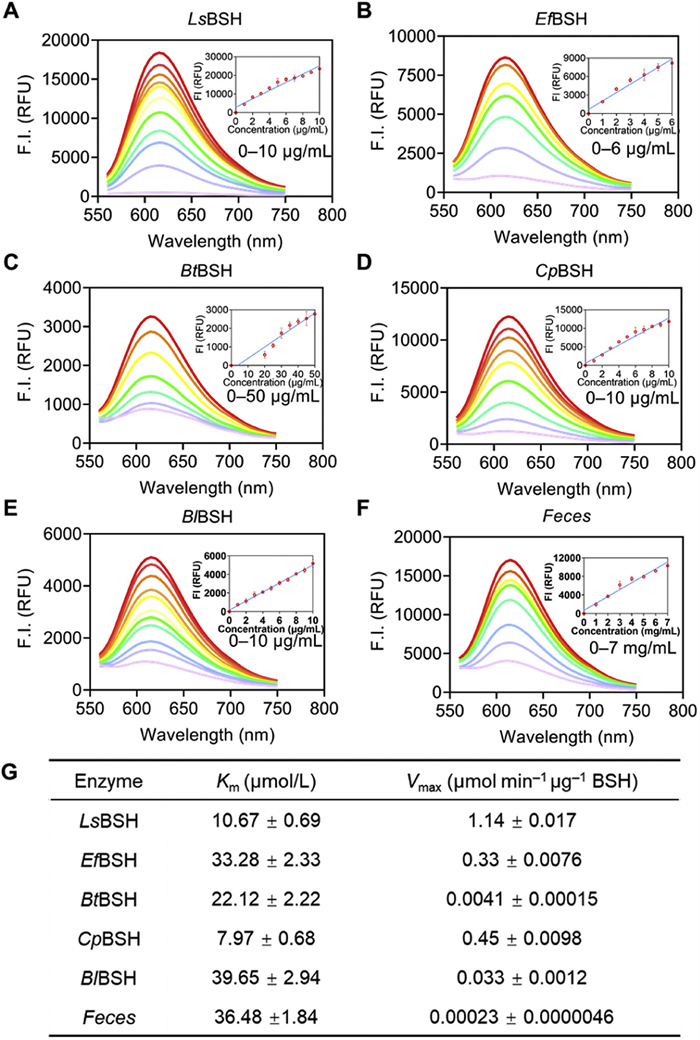
We further characterized the sensing capabilities of CA-ABEI by determining its linear detection range and sensitivity toward BSH. As shown in Fig. S7 (Supporting information), the fluorescence intensity of CA-ABEI demonstrated excellent linear correlation with reaction time during a specific incubation period, revealing its time-dependent sensing capability. Furthermore, the quantitative relationship between fluorescence signal and recombinant BSH concentration was systematically examined. Figs. 3A–F demonstrated a concentration-dependent enhancement of fluorescence intensity, revealing a well-defined linear correlation within the 0–10 µg/mL (LsBSH, R2 = 0.94), 0–6 µg/mL (EfBSH, R2 = 0.95), 0–50 µg/mL (BtBSH, R2 = 0.92), 0–10 µg/mL (CpBSH, R2 = 0.96), 0–10 µg/mL (BlBSH, R2 = 0.98) and 0–7 mg/mL (Feces, R2 = 0.95). The probe exhibited linear relationships with respect to both reaction time and BSH concentration, enabling precise enzyme activity measurement and kinetic investigations. Enzyme kinetics is critical for assessing catalytic activity and identifying potential inhibitors for specific enzymatic targets [33]. The hydrolysis kinetics of CA-ABEI towards BSH was carefully characterized. As shown in Fig. 3G and Fig. S9 (Supporting information), the hydrolytic reaction of CA-ABEI towards BSHs followed the classic Michaelis–Menten kinetics. The Km of CA-ABEI in BSH were 10.67 µmol/L (LsBSH), 33.28 µmol/L (EfBSH), 22.12 µmol/L (BtBSH), 7.97 µmol/L (CpBSH), 39.65 µmol/L (BlBSH) and 36.48 µmol/L (Feces). LsBSH was showing high affinity and rapid hydrolytic rate. The detection sensitivity of CA-ABEI was further assessed across various BSHs, yielding LODs of 0.06 µg/mL (LsBSH), 0.08 µg/mL (EfBSH), 1.04 µg/mL (BtBSH), 0.33 µg/mL (CpBSH), 0.21 µg/mL (BlBSH), and 240 µg/mL (mouse feces) (Table S3 in Supporting information). The investigation demonstrated that CA-ABEI is an efficient and reliable fluorogenic substrate for BSH, exhibiting excellent performance in both enzyme activity assays and practical applications involving mammalian fecal samples.
Figure 3
Figure 3.
The fluorescence spectra of CA-ABEI under LsBSH (A), EfBSH (B), BtBSH (C), CpBSH (D), BlBSH (E) and Feces (F). Enzyme kinetic parameters of CA-ABEI under different enzyme sources (G). λex = 520 nm, λem = 620 nm. Data are presented as mean ± SD (n = 3).
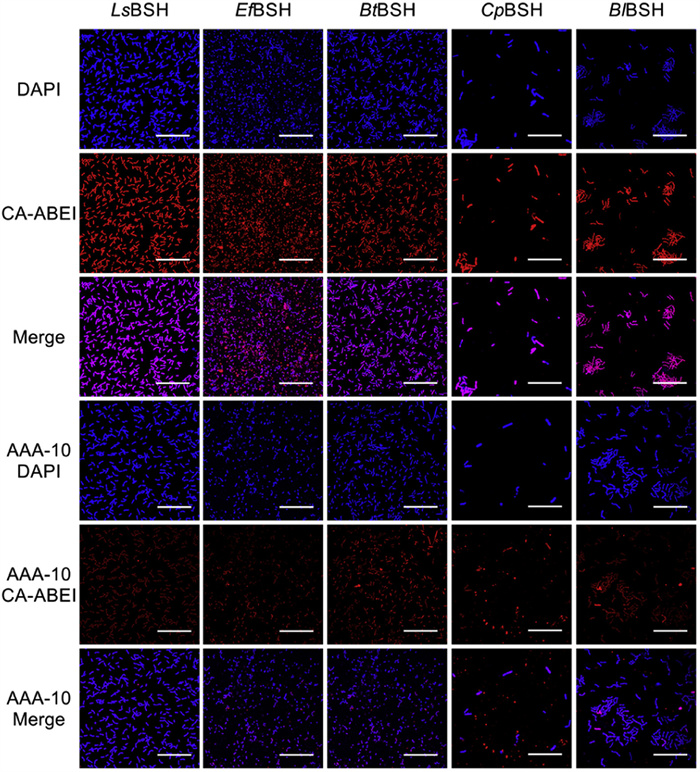
In recent years, fluorescent probes have been widely applied in medical imaging due to their advantages such as low cost, high specificity, non-invasiveness, high resolution, and superior signal-to-noise ratio [34–36]. The high specificity and long emission wavelength of the fluorogenic substrate enable the detection of BSH activity in live bacteria during bioimaging studies. Following a 1 h incubation at 37 ℃ with BSH-expressing E. coli, CA-ABEI generated a distinct red fluorescence signal detectable by confocal microscopy (Fig. S10 in Supporting information). The marked reduction in fluorescence intensity upon treatment with AAA-10, a selective BSH inhibitor, confirmed the probe's capability for endogenous BSH detection in live bacterial systems. Among E. coli strains specifically expressing different BSH, the fluorescence signal in the red channel was highest for LsBSH, followed by EfBSH, CpBSH, BlBSH, and BtBSH, which were consistent with the fluorescence intensity obtained from in vitro enzyme assays (Fig. S11 in Supporting information), thereby reinforcing the reliability of CA-ABEI as a practical fluorogenic probe of BSH. Inspired by the above data, CA-ABEI was used to specifically detect the BSH activity within these five natural intestinal microbiota strains. As depicted (Fig. 4), CA-ABEI accurately detected the BSH activity within the bacteria without altering their morphology. These results strongly confirmed that CA-ABEI exhibited excellent biocompatibility and could efficiently and specifically detect BSH in live bacteria, thereby providing strong evidence for its application potential in microbiological research and related fields.
Figure 4
Figure 4.
Confocal fluorescence images of the natural intestinal microbiota expressing the five specific bsh genes were incubated with CA-ABEI (20 µmol/L) at 37 ℃ for 1 h in the presence and absence of AAA-10 (200 µmol/L). λex = 488 nm, λem = 610–630 nm (100 × oil objective). Scale bar: 25 µm.
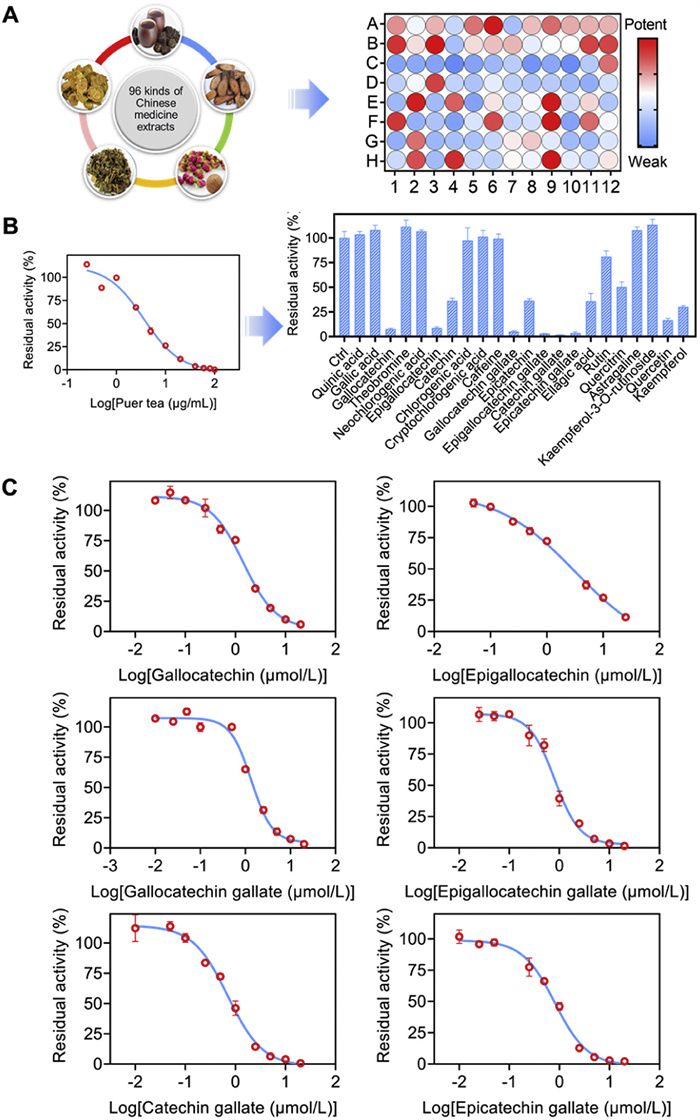
A multitude of studies have demonstrated that inhibitors of BSH can attain the effect of treating diseases by altering the bile acid profile within the host's intestine [37–39]. In this study, we utilized CA-ABEI to establish a novel fluorescence detection platform, facilitating rapid and effective high-throughput screening of potential BSH inhibitors. AAA-10, a known positive inhibitor of BSH, demonstrated dose-dependent inhibition of BSH activity, with an IC50 value consistent with previously reported data (Fig. S12 in Supporting information) [40]. The aforementioned results fully validated the efficacy of the newly developed method. Given the unique advantages of natural compounds, such as chemical diversity, multifunctionality and low toxicity, there has been increasing interest in exploring traditional Chinese medicine for the development of novel enzyme inhibitors. Consequently, 96 traditional Chinese medicine extracts were screened for their inhibitory effects on LsBSH using CA-ABEI as the fluorogenic substrate (Fig. 5A and Table S4 in Supporting information). Among the tested extracts, six traditional Chinese medicines, including Rheum palmatum L., Polygonum multiflorum Thunb., Agrimonia pilosa Ledeb., Fallopia multiflora (Thunb.) Harald., Rosa rugosa Thunb., and Pu-erh tea exhibited significantly inhibitory activities against LsBSH (Table S5 in Supporting information). Pu-erh tea exhibited the strongest inhibition of LsBSH activity, showing an IC50 of 3.61 ± 0.35 µg/mL (Fig. 5B). To identify the active components responsible for this effect, the inhibitory potency of 22 major previously reported compounds in Pu-erh tea was further evaluated (Table S6 in Supporting information) [41]. Among them, gallocatechin, epigallocatechin, gallocatechin gallate, epigallocatechin gallate, catechin gallate, and epicatechin gallate exhibited significant inhibitory effects against LsBSH, with IC50 values of 1.44, 3.18, 1.35, 0.81, 0.71 and 0.82 µmol/L, respectively (Fig. 5C and Table S7 in Supporting information). These results indicated that the CA-ABEI-based fluorescent assay represents a reliable and effective methodology for high-throughput screening and evaluation of BSH inhibitors derived from traditional Chinese medicinal sources and their chemical constituents.
Figure 5
Figure 5.
High-throughput screening of BSH inhibitors. (A) Screening BSH inhibitors from 96 kinds of traditional Chinese medicine extracts (100 µg/mL) using CA-ABEI (10 µmol/L) as the substrate. (B) Determination of the inhibitory effect of the components (10 µmol/L) in Pu-erh tea against LsBSH (2 µg/mL). (C) The IC50 values of several compounds in Pu-erh tea against LsBSH (2 µg/mL). λex = 520 nm, λem = 620 nm. Data are presented as mean ± SD (n = 3).
In summary, a universal far-red fluorogenic substrate, CA-ABEI, was successfully developed from aminocoumarin fluorophore and was specifically recognized by BSH. CA-ABEI exhibited excellent fluorescent properties, a favorable signal-to-noise ratio, high stability, specificity, and outstanding biocompatibility. Notably, CA-ABEI enabled the first specific fluorescent imaging of BSH in live bacteria with high resolution. Furthermore, the CA-ABEI-based detection system was applied to establish a high-throughput screening platform for identifying potential BSH inhibitors. Among 96 traditional Chinese medicine extracts, Pu-erh tea was identified as a potent BSH inhibitor and its key active components were further analyzed, providing a strong foundation for investigating the inhibitory effects of Pu-erh against BSH. Collectively, CA-ABEI proved to be a powerful tool for monitoring BSH activity in complex biological systems with value for exploring physiological functions and rapid screening of inhibitors.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
This study was supported by the National Natural Science Foundation of China (Nos. 82274011, U23A20516, U24A20793 and 82260843), the National Key Research and Development Program of China (No. 2022YFC3502000), and the Traditional Chinese Medicine Guangdong Provincial Laboratory Scientific Research and Development Incubation Project (No. HQL2024PZ004).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2025.111320.
C. Xie, C.T. Jiang, J.M. Shi, et al., Diabetes 66 (2017) 613–626. doi: 10.2337/db16-0663
[40]
A.A. Adhikari, D. Ramachandran, S.N. Chaudhari, et al., ACS Chem. Biol. 16 (2021) 1401–1412. doi: 10.1021/acschembio.1c00192
[41]
Z.Q. Zhou, Y. Li, F.Y. Wang, et al., Food Funct. 15 (2024) 5287–5299. doi: 10.1039/d4fo00538d
Figure 1
Rational design and screening of fluorogenic substrates for BSHs. (A) Design and virtual screening of BSH fluorogenic substrates. (B) The fluorescence response of S2–S8 (10 µmol/L) after reaction with five BSHs (20 µg/mL). (C) Fluorescence spectral changes of CA-ABEI (10 µmol/L) after reaction with five BSHs (20 µg/mL). λem = 520 nm, data are presented as mean ± SD (n = 3).
Figure 2
The sensing mechanism and enzyme specificity of CA-ABEI towards BSHs. (A) The sensing mechanism of CA-ABEI for BSH. (B) HPLC characterization of BSH—catalyzed hydrolysis of CA-ABEI. (C) The selectivity of CA-ABEI (10 µmol/L) among various hydrolases (10 µg/mL). λex = 520 nm, λem = 620 nm. Data are presented as mean ± SD (n = 3).
Figure 3
The fluorescence spectra of CA-ABEI under LsBSH (A), EfBSH (B), BtBSH (C), CpBSH (D), BlBSH (E) and Feces (F). Enzyme kinetic parameters of CA-ABEI under different enzyme sources (G). λex = 520 nm, λem = 620 nm. Data are presented as mean ± SD (n = 3).
Figure 4
Confocal fluorescence images of the natural intestinal microbiota expressing the five specific bsh genes were incubated with CA-ABEI (20 µmol/L) at 37 ℃ for 1 h in the presence and absence of AAA-10 (200 µmol/L). λex = 488 nm, λem = 610–630 nm (100 × oil objective). Scale bar: 25 µm.
Figure 5
High-throughput screening of BSH inhibitors. (A) Screening BSH inhibitors from 96 kinds of traditional Chinese medicine extracts (100 µg/mL) using CA-ABEI (10 µmol/L) as the substrate. (B) Determination of the inhibitory effect of the components (10 µmol/L) in Pu-erh tea against LsBSH (2 µg/mL). (C) The IC50 values of several compounds in Pu-erh tea against LsBSH (2 µg/mL). λex = 520 nm, λem = 620 nm. Data are presented as mean ± SD (n = 3).

 DownLoad:
DownLoad:

 下载:
下载:




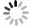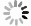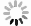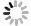

 下载:
下载:
