State Key Laboratory of Analytical Chemistry for Life Science, School of Chemistry and Chemical Engineering, Nanjing University, Nanjing 210023, China
b.
Department of Reproductive Medical Center, Jinling Hospital, School of Medicine, Nanjing University, Nanjing 210002, China
c.
Chemistry and Biomedicine Innovation Center (ChemBIC), Nanjing University, Nanjing 210023, China
d.
Center of Clinical Laboratory Science, The Affiliated Cancer Hospital of Nanjing Medical University & Jiangsu Cancer Hospital & Jiangsu Institute of Cancer Research, Nanjing 210009, China
* Corresponding authors at: Department of Reproductive Medical Center, Jinling Hospital, School of Medicine, Nanjing University, Nanjing 210002, China. ** Corresponding author at: State Key Laboratory of Analytical Chemistry for Life Science, School of Chemistry and Chemical Engineering, Nanjing University, Nanjing 210023, China. E-mail addresses: zhaowei0714@126.com (W. Zhao), yaobing@nju.edu.cn (B. Yao), dinglin@nju.edu.cn (L. Ding).
Received Date:
7 March 2023 Revised Date:
8 June 2023 Accepted Date:
30 June 2023 Available Online:
1 July 2023
Spermatogenesis, maturation, capacitation and fertilization are precisely regulated by glycosylation. However, the relationship between altered glycosylation patterns and the onset and development of reproductive disorders is unclear, mainly limited by the lack of in situ imaging techniques for spermatozoa glycosylation. We developed an efficient and highly specific spermatozoa glycan imaging technique based on the robust chemoselective labeling of sialic acid (Sia) and N-acetyl-d-galactosamine (Gal/GalNAc). We further proposed a "tandem glycan chemoselective labeling" strategy to achieve simultaneous imaging of two types of glycans on spermatozoa. We applied the developed method to the spermatozoa from oligozoospermic patients and diabetic mice and found that these spermatozoa showed higher levels of Sia and Gal/GalNAc expression than the normal groups. Moreover, spermatozoa from diabetic mice showed a severe decrease in number, viability, and forward motility, suggesting that in vivo glucose metabolism disorders may lead to an elevated level of spermatozoa glycosylation and have a correlation with the development of oligoasthenotspermia. Our work provides a research tool to reveal the relationship between glycosylation modification and spermatozoa quality, and a promising clue for the development of glycan-based reproductive markers.
The process of spermatogenesis in mammals is complex and sophisticated. Spermatogonia go through mitotic, meiotic, and post-meiotic processes in the testis before eventually forming spermatozoa [1]. After entering the epididymis, the immature spermatozoa undergo a magnificent structural and functional transformation as they interact with the surrounding epididymal media in preparation for fertilization [2]. In clinical, the quality of spermatozoa is generally evaluated by measuring their physiological parameters such as quantity, viability, and morphology in the semen [3]. However, these parameters are still far from being sufficient to reveal the regulatory factors of the spermatogenesis and development process, as well as to elucidate the pathophysiology of oligozoospermia, asthenozoospermia, and teratozoospermia caused by common reproductive diseases [4,5]. Hence, there is an urgent need to develop markers at the molecular level, which is of great importance for the development of assisted reproduction techniques to improve spermatozoa quality [6,7].
The process of spermatogenesis, maturation, and fertilization is precisely regulated by proteins [7–9]. In particular, the post-translational modification (PTM) status of proteins, including glycosylation, phosphorylation, acetylation, nitrosylation, and methylation, etc., is a key factor in determining spermatozoa production, maturation, and ultimately spermatozoa-egg binding [10–12]. Among them, glycosylation modifications are the most diverse, with complex glycan sequences regulating a wide range of physiological functions in spermatozoa [13–15].
Glycosylation is the process of attaching glycans to proteins or lipids to form covalent bonds under the control of glycan-related enzymes [16–20]. Up to 92% of proteins on the outer membrane of mammalian spermatozoa are glycoproteins [15]. Glycans themselves build a protective barrier for spermatozoa and enhance the communication between spermatozoa and external media [21]. Glycosylation on the spermatozoa surface also plays an important role in the process of spermatozoa-egg binding [22–27]. A few studies have found that the development of reproductive diseases is accompanied by altered glycosylation levels. In the semen of patients with leukocytospermia, the fucosylation of fibronectin is reduced, while its sialylation is increased, similarly, the sialylation of IgA secretory component (SC) glycoprotein is also increased [28,29]. In the seminal plasma of infertile oligozoospermic patients, the abundance of glycoprotein lactotransferrin is more prevalent [30]. Therefore, glycosylation is a promising biomarker that can help not only to elucidate the molecular mechanisms of the fertility process and to discover the etiology of male reproductive diseases, but also to develop new methods for the diagnosis and intervention of clinical androgenic defects [31]. However, studies on glycan expression in spermatozoa of oligoasthenotspermia models are largely lacking.
Difficulties in developing glycan biomarkers for reproductive research, detection, and intervention lie in (1) the complexity of glycan labeling and detection, and (2) the specificity of spermatozoa. Unlike nucleic acids and proteins, glycans are not a direct product of genetic coding [32]. Because spermatozoa lose their ability to manufacture glycans once they leave the testis, it is challenging to label spermatozoa in vitro using the classical metabolic oligosaccharide engineering (MOE) approaches [14,32]. Currently, there are two main types of methods for spermatozoa glycan analysis: (1) glycomics techniques and (2) techniques based on lectin recognition of glycans (lectin fluorescent probes [33], lectin microarrays [34]). The most effective method for revealing glycan structure is mass spectrometry-based glycomics technology. Pang group [26] examined the N-glycome of normal human spermatozoa by MALDI-TOF MS and found the acrosome region of spermatozoa contained Lewisx and Lewisy modified glycoproteins. 297 N-glycosylated proteins were found in human spermatozoa by the Guo group [35]. Sun group [36] investigated the glycan structure at various glycosylation sites on human spermatozoa glycoproteins. However, mass spectrometry is a destructive technique that cannot reflect the distribution of glycans on spermatozoa in situ. Spermatozoa are terminal cells that cannot divide and proliferate by themselves in vitro [37], and their in vitro culture is also very difficult, so generally they should be taken and measured immediately to insure their intrinsic nature. Lectins are non-immunogenic proteins that can specifically recognize glycans and distinguish among glycoforms and thus could be used for in situ imaging of spermatozoa [33]. However, lectins have poor specificity and low affinity, and it is difficult to exclude non-specific signals of lectin adsorption from the results. Moreover, their non-covalent binding is easily dissociated by interference from external factors; and lectins generally contain multiple recognition subunits, leading to cell aggregation that interferes with the assay results. To in situ uncover the expression information of glycans in aberrant mammalian spermatozoa, a sensitive, specific, and efficient spermatozoa glycan detection technology is urgently needed, which can provide powerful tools for the development of glycan-based reproductive markers [14].
Herein, we used Chemoselective Glycan Labeling (CGL) strategy to establish a novel method for in situ fluorescence imaging and quantitative flow cytometric detection of spermatozoa glycans. CGL has the advantages of being performed from outside of the cell (independent of cellular physiological function), short labeling time (~min), high specificity, and the formation of bioorthogonal covalent chemical labeling [38,39]. We used sodium periodate (NaIO4) and galactose oxidase (GAO) to selectively oxidize terminal sialic acid (Sia) and galactose/N-acetyl-d-galactosamine (Gal/GalNAc) to generate aldehyde groups, respectively (Fig. 1a). Combined with click chemistry and biotin-streptavidin recognition, robust Sia and Gal/GalNAc labeling on spermatozoa was achieved in situ. Based on this, we designed a "tandem glycan chemoselective labeling" strategy to label Sia and Gal/GalNAc on spermatozoa, allowing the acquisition of in situ distribution information of both glycans on a single spermatozoon simultaneously. Importantly, we applied the method to study the glycosylation levels in a model of oligoasthenotspermia and found that the expression of Sia and Gal/GalNAc on spermatozoa were significantly higher in the oligoasthenotspermia group than those of the normal group. Further, considering the correlation between glucose metabolism disorders and infertility phenotypes, we used diabetic mice (db/db) as a model and found their spermatozoa had abnormally higher levels of glycan expression, which correlated with increased defectivity of spermatozoa.
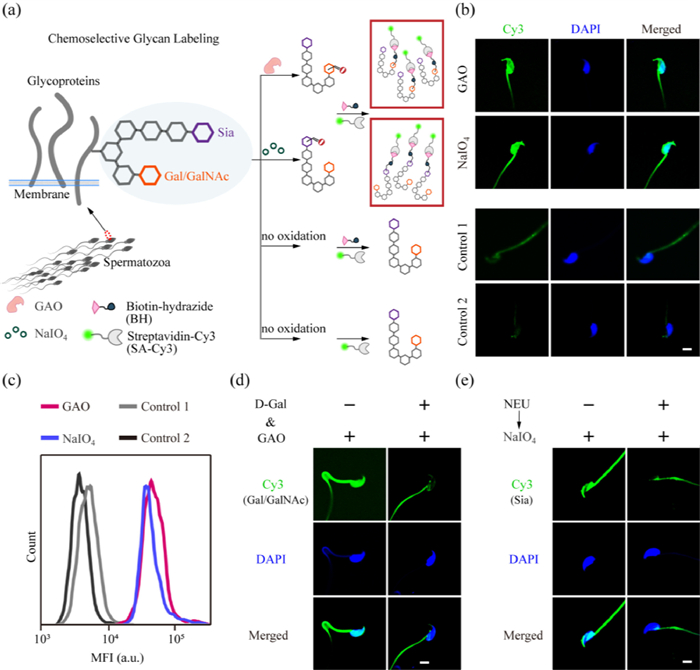
Figure 1
Figure 1.
(a) Schematic illustration of the Chemoselective Glycan Labeling (CGL) strategy. (b) Confocal laser scanning microscopy (CLSM) images of mice spermatozoa after GAO or NaIO4 oxidation, biotin-hydrazide (BH) linking, and streptavidin-Cy3 (SA-Cy3) binding. Control 1: spermatozoa treated with BH and then SA-Cy3 (without oxidation). Control 2: spermatozoa treated only with SA-Cy3. (c) Flow cytometric analysis of spermatozoa in (b). (d) CLSM images of spermatozoa undergoing GAO oxidation in the presence or absence of d-Gal, and BH→SA-Cy3 labeling. (e) CLSM images of spermatozoa with or without pretreatment by neuraminidase (NEU) followed by NaIO4 oxidation and BH→SA-Cy3 labeling. Cell nuclei were stained with DAPI (blue). Scale bars: 5 µm. The above data are representative of three individual experiments.
The glycan glycoforms specific to pathogens and cancer cells, although rarely present in healthy human body fluids, are abundant on the spermatozoa surface as well as in seminal plasma glycoproteins of healthy fertile individuals [40]. This may be because spermatozoa exploit the same glycoforms as tumor cells to evade killing by immune cells [26]. Based on these similarities, we first used easily accessible and cultured tumor cells (MCF-7 (human breast cancer cell line), HeLa (human cervical cancer cell line), HepG2 (human liver cancer cell line)) as a model to demonstrate the oxidation labeling of Sia and Gal/GalNAc on adhered cells in confocal dishes. The key to selective labeling of Sia lies in the control of the oxidation capacity of NaIO4 [41]. We treated the cells with NaIO4 solution (2 mmol/L) at 4 ℃ to oxidize the cis‑hydroxyl groups at the C-7 and C-8 positions of Sia to generate aldehyde groups; while for Gal/GalNAc, we incubated the cells with GAO to oxidize the hydroxyl groups at the C-6 position of Gal/GalNAc to generate aldehyde groups [42]. Aldehyde groups are bio-orthogonal groups on the cell surface and can react with hydrazide-modified labeling molecules (e.g., fluorescent molecules, biotin) to create hydrazone linkage under the catalysis of aniline. We successfully achieved the chemoselective covalent labeling of Sia and Gal/GalNAc on tumor cells using fluorescein-5-thiosemicarbazide (FTZ) as a fluorescent probe (Figs. S1–S4 in Supporting information).
We then proceeded to establish a fluorescence imaging method for mouse spermatozoa glycans based on CGL. We used the 12-week-old C57BL/6 adult male mice as a model. All experimental protocols were approved by the Institutional Animal Care and Use Committee of Nanjing Jinling Hospital (2020JLHGKJDWLS-169). The mice caudal epididymides were removed and crushed, filtered through a cell strainer, centrifuged, and then washed with PBS to obtain spermatozoa samples. To prevent alterations in the glycans of spermatozoa caused by changes in the external environment after isolation, we fixed and blocked the spermatozoa immediately after separation. To achieve the imaging and flow cytometric analysis of mouse spermatozoa, we used biotin-hydrazide (BH) to react with the aldehyde groups generated by NaIO4/GAO oxidation on mouse spermatozoa, followed by streptavidin-Cy3 (SA-Cy3) staining (Fig. 1a). We tried various hydrazide labeling reagents and found that SA-Cy3 combined with BH had the lowest adsorption background for labeling of aldehyde groups generated on spermatozoa. Unlike the glycan imaging results for live tumor cells in Figs. S1–S4, the whole fixed spermatozoa showed fluorescence signals (Fig. 1b). It should be noted that because spermatozoa are highly asymmetrical cells, when we focus on the head of the spermatozoa using DAPI, the tail may or may not be in the plane of focus. In the group without oxidation (Control 1, BH and then SA-Cy3), the fluorescence was substantially diminished, indicating that the fluorescence observed in the experimental group mainly originated from the selective labeling of glycans. In addition, in group Control 2 (only SA-Cy3), the background fluorescence of spermatozoa was further attenuated, indicating that the background fluorescence originated mainly from the adsorption of BH. We also performed flow cytometric analysis of the labeled spermatozoa and found that the signal-to-background ratio of GAO group to Control 1 was about 9.21, and the ratio for the NaIO4 group to Control 1 was about 7.93 (Fig. 1c).
Next, we verified the specificity of Gal/GalNAc and Sia labeling using monosaccharide competition assay and neuraminidase (NEU) cleavage experiment, respectively. When free d-(+)-galactose (d-Gal) was added to compete with spermatozoa for GAO oxidation, a decrease in fluorescence intensity on spermatozoa was observed, demonstrating that the oxidation target of GAO is indeed Gal/GalNAc (Fig. 1d). On the other hand, NEU can specifically cleave α2–3,6,8 linked Sia. The pretreatment of spermatozoa by NEU prior to NaIO4 oxidation step led to substantially reduced fluorescence signals, demonstrating the specificity of Sia labeling (Fig. 1e).
We validated the results of the chemoselective labeling methods using the traditional lectin recognition technique. Five types of lectins (FITC labeled) were used: wheat germ agglutinin (WGA), Sambucus nigra agglutinin (SNA), Jacalin, peanut agglutinin (PNA), soybean agglutinin (SBA). The detailed recognition information was shown in Table S1 (Supporting information). WGA, SNA, and Jacalin showed strong binding to spermatozoa, the binding of PNA was moderate, while the binding of SBA was negligible (Fig. S5 in Supporting information). These results were consistent with our results, thus demonstrating the reliability of chemoselective labeling.
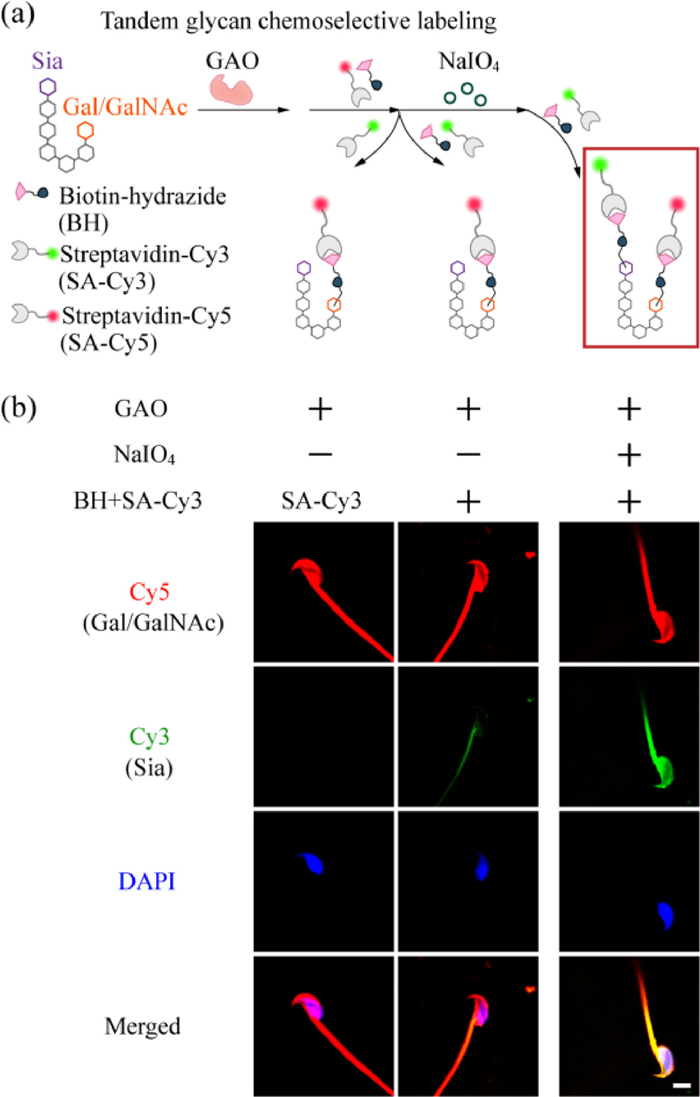
We then attempted to simultaneously image two types of glycans (Sia and Gal/GalNAc), on the same spermatozoon, so that we could assess the expression pattern of glycans on spermatozoa at the "single cell" level. We designed a new method called "tandem glycan chemoselective labeling", which consists of two main steps: (1) GAO oxidation of terminal Gal/GalNAc on spermatozoa, followed by sequential attachment of BH and streptavidin-Cy5 (SA-Cy5), and (2) blocking of excess aldehyde groups, and then NaIO4 oxidation of Sia on spermatozoa, followed by sequential attachment of BH and SA-Cy3 (Fig. 2a). After labeling was completed, the nuclei were indicated with DAPI and CLSM imaging was performed (Fig. 2b). In the experimental group that the full set of labeling was performed, red (Cy5) and green (Cy3) fluorescence indicated Gal/GalNAc and Sia, respectively (right); whereas in the group without NaIO4 treatment (middle), only SA-Cy5 fluorescence from GAO-mediated labeling was observed.
Figure 2
Figure 2.
(a) Schematic illustration of the tandem glycan chemoselective labeling technique. (b) CLSM images of spermatozoa after different treatment combinations shown in (a). Cell nuclei were stained with DAPI (blue). Scale bar: 5 µm. All images are representative of three individual experiments.
According to the literature, oligoasthenotspermia is one of the important causes of male infertility [4,8]. Kratz group [43] compared the expression of glycans in the seminal plasma of normal and oligozoospermic patients using a series of lectins and found that the ability of glycans to bind SNA was significantly lower in the seminal plasma of oligozoospermic patients compared to the normal group; the glycosylation levels of several glycoproteins (α‐1‐acid glycoprotein (AGP), fibronectin (FN), immunoglobulin G (IgG), glycodelin S (GdS), etc.) are also dysregulated in some seminal plasma samples with abnormal semen parameters [31]. Maturation of spermatozoa in the epididymis depends on the interaction with various enzymes and substrates in the seminal fluid [2,44], and changes in the microenvironment in the seminal fluid could largely lead to alterations in the modifications of spermatozoa, so we speculated that in oligoasthenotspermia model, the glycans of spermatozoa may also be altered.
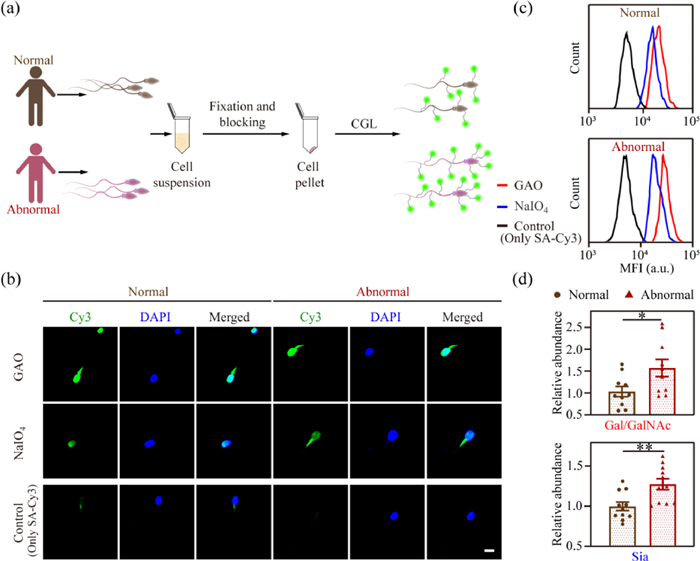
To verify this hypothesis, we collected 10 groups of semen from oligozoospermic patients and 10 groups from healthy people, and extracted the spermatozoa (Fig. 3a). All the experiments were approved by the Research Ethics Committee of Nanjing Jinling Hospital (2020DZGZRZX-094). We first labeled Gal/GalNAc and Sia of the spermatozoa, and used CLSM imaging to verify the feasibility of the method for human spermatozoa labeling. We found that the fluorescence intensity of glycans on the spermatozoa of the two labeled groups was significantly higher than that of the control group (Fig. 3b), and the spermatozoa morphology did not change significantly.
Figure 3
Figure 3.
(a) Schematic showing the labeling protocol of spermatozoa from clinical normal and abnormal human semen samples. (b) Representative CLSM images and (c) flow cytometric results of normal and abnormal human spermatozoa undergoing CGL and fluorescence staining. Scale bar: 5 µm. (d) Quantification of median fluorescence intensity (MFI) in (c). The expression level of glycans (Gal/GalNAc or Sia) in abnormal spermatozoa relative to that of normal spermatozoa was shown. Data are shown as the mean ± SEM, n = 10 (t-test). P < 0.05; **P < 0.01.
Further, we quantified glycan expression using flow cytometric analysis (Fig. 3c). We found that in both normal group and oligoasthenotspermia group, the fluorescence signal of Gal/GalNAc was always higher than that of Sia, however, the expression of Gal/GalNAc and Sia increased by 69.8% and 28.5%, respectively, in oligoasthenotspermia group compared with the normal group, with the increase of Gal/GalNAc more obvious (Fig. 3d). Agarwal group [45] reported that the occurrence of oligoasthenotspermia is associated with an increase in spermatozoa defectivity, and Pang group found that defective spermatozoa generally show high levels of Lewisy glycoform expression [26], resulting in more Gal/GalNAc exposed. Therefore, we hypothesized that the rise in terminal glycan Gal/GalNAc we observed might be partially caused by enhanced Lewisy expression.
Diabetes mellitus (DM) is proved to be one of the causative factors of oligoasthenotspermia. As a serious disorder of glucose metabolism, DM also leads to increased levels of glycosylation [13,46]. Therefore, we speculated that the glycan expression of spermatozoa in diabetic models might also be altered.
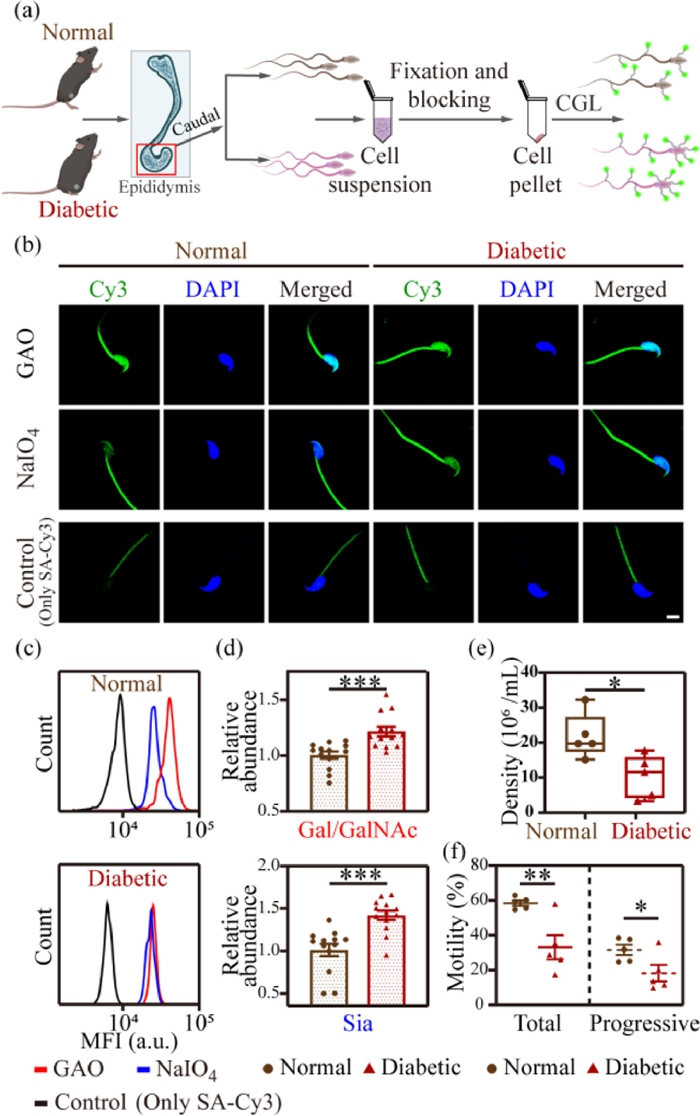
To test this, we first extracted spermatozoa from the caudal epididymis of diabetic (db/db) mice (experimental group) and normal (BKS) mice (control group), respectively (Fig. 4a), and examined the expression of Gal/GalNAc and Sia using CLSM imaging (Fig. 4b) and flow cytometric analysis (Fig. 4c). We found that the expression of Gal/GalNAc and Sia on the spermatozoa of db/db mice was 21.4% and 44.4% higher than that of BKS mice, respectively (Fig. 4d). Unlike the variation pattern in oligozoospermic patients, the alteration trend for Sia on spermatozoa was more pronounced in db/db mice. According to the literature, DM has a serious impact on the reproductive system, leading to increased spermatozoa DNA damage and decreased motility [47–49]. We then extracted spermatozoa from the caudal epididymis of five db/db mice groups and tested their quality, including spermatozoa count and viability (Figs. 4e and f). The spermatozoa density in the caudal epididymis of db/db mice was 10.74 × 106/mL, compared with 22.43 × 106/mL in BKS mice. Additionally, the spermatozoa motility rate of db/db mice was 32.28%, which was significantly lower than that of BKS mice (57.97%); the progressive motility rate was 17.23%, which was also significantly lower than that of the control group (30.74%).
Figure 4
Figure 4.
(a) Schematic showing the labeling protocol of spermatozoa from normal (BKS) and diabetic (db/db) mice. (b) Representative CLSM images and (c) flow cytometric results of BKS and db/db mice spermatozoa undergoing CGL labeling and fluorescence staining. Scale bar: 5 µm. (d) Quantification of MFI in (c), n = 13 (t-test). The expression level of glycans (Gal/GalNAc or Sia) in spermatozoa from diabetic mice relative to that from normal mice was shown. (e) Spermatozoa density of BKS mice and db/db mice (n = 5 (t-test)) are shown as points in the boxplots, the center lines represent the median, box edges show the 25th and 75th percentiles, and whiskers represent min to max. (f) Motility detection of spermatozoa from BKS mice and db/db mice, n = 5 (t-test). Data in (d) and (f) are shown as the mean ± SEM. P < 0.05; **P < 0.01; ***P < 0.001.
The above results indicated that the quantity and motility of spermatozoa in diabetic mice were severely decreased, and this increase in spermatozoa defectivity was accompanied by an increase in Sia and Gal/GalNAc expression. These validate our speculation that glucose metabolism disorders in vivo may lead to increased levels of spermatozoa glycosylation, which is tightly correlated with the occurrence of oligoasthenotspermia.
In conclusion, we developed a technique for in situ imaging and quantitative analysis of mouse and human spermatozoa glycans based on Chemoselective Glycan Labeling. The simultaneous imaging of both glycans on a single spermatozoon was achieved by developing a tandem glycan chemoselective labeling method. We used the developed method for imaging and detection of two important glycans (Sia and Gal/GalNAc) on spermatozoa from oligozoospermic patients and found elevated levels of sialylation and terminal Gal/GalNAc glycosylation in spermatozoa from the oligoasthenotspermia group. This serves as a crucial resource for future research into the molecular mechanisms of spermatozoa motility and other functions as well as the development of clinically targeted therapies for reproductive diseases. Taking this as a clue, we further conducted in situ studies of spermatozoa glycosylation in a diabetic model. As a disorder of glucose metabolism in the organism, diabetes generally causes a decrease in spermatozoa quality, which can severely interfere with reproductive function. Many reproduction-related proteins exhibit up- and down-regulated expression in diabetic patients [47], and more importantly, their glycosylation modifications are significantly altered, which are closely related to spermatozoa motility, acrosome response, and fertilization ability. The elevated glucose level [19] in diabetic patients may lead to increased levels of O-GlcNAc glycosylation of proteins, in addition, altered glycosylation of reproduction-related luteinizing hormone (LH) has also been detected [50]. We found elevated levels of Sia and terminal Gal/GalNAc expression in spermatozoa of diabetic mice, and we speculated that the increased glycan expression may affect protein half-life, accelerate cellular oxidative processes, and lead to sustained release of ROS and impaired mitochondrial function, which ultimately alters the morphology and activity of spermatozoa, causing a decrease in their quality [51,52]. Currently, several biomarkers of glycosylation have been shown to play a role in the clinical diagnosis of male infertility, such as the sialylated Lewisx antigen of α−1-acid glycoprotein [28], which is significantly increased in chronic inflammation of the male reproductive tract (also known as leukocytospermia). Our work provides a new perspective to study the molecular mechanisms of male infertility; a research tool for revealing the relevance between glycosylation modification and spermatozoa quality; and potential glycan-based biomarkers for clinical diagnosis of male infertility caused by spermatozoa defect problems. We believe that our research establishes a solid foundation for the development of the next stage of glycan modification-based reproductive intervention technologies.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We gratefully acknowledge the support from the National Natural Science Foundation of China (Nos. 21974067, 22274073, 81971373 and 82001535), the National Key Research and Development Program of China (No. 2018YFC1004700), Fundamental Research Funds for the Central Universities (Nos. 020514380309, 021414380502 and 2022300324), and the State Key Laboratory of Analytical Chemistry for Life Science (Nos. 5431ZZXM2305 and 5431ZZXM2204).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2023.108760.
Jiajia Lv
, Jie Gao
, Hongyu Li
, Zeli Yuan
, Nan Dong
. Rational design of hydroxytricyanopyrrole-based probes with high affinity and rapid visualization for amyloid-β aggregates in vitro and in vivo. Chinese Chemical Letters,
2024, 35(5): 108940-.
doi: 10.1016/j.cclet.2023.108940
Sixin Ai
, Wenxiu Li
, Huayong Zhu
, Yang Wan
, Weiying Lin
. Viscosity-responsive signal amplification dual-modal probe triggered by cysteine/homocysteine for monitoring diabetic liver damages and repair processes. Chinese Chemical Letters,
2025, 36(3): 109904-.
doi: 10.1016/j.cclet.2024.109904
Yiwen Lin
, Yijie Chen
, Chunhui Deng
, Nianrong Sun
. Integration of resol/block-copolymer carbonization and machine learning: A convenient approach for precise monitoring of glycan-associated disorders. Chinese Chemical Letters,
2024, 35(12): 109813-.
doi: 10.1016/j.cclet.2024.109813
Linfang ZHANG
, Wenzhu YIN
, Gui YIN
. A 2-dicyanomethylene-3-cyano-4,5,5-trimethyl-2,5-dihydrofuran-based near-infrared fluorescence probe for the detection of hydrogen sulfide and imaging of living cells. Chinese Journal of Inorganic Chemistry,
2025, 41(3): 540-548.
doi: 10.11862/CJIC.20240405
Figure 1. (a) Schematic illustration of the Chemoselective Glycan Labeling (CGL) strategy. (b) Confocal laser scanning microscopy (CLSM) images of mice spermatozoa after GAO or NaIO4 oxidation, biotin-hydrazide (BH) linking, and streptavidin-Cy3 (SA-Cy3) binding. Control 1: spermatozoa treated with BH and then SA-Cy3 (without oxidation). Control 2: spermatozoa treated only with SA-Cy3. (c) Flow cytometric analysis of spermatozoa in (b). (d) CLSM images of spermatozoa undergoing GAO oxidation in the presence or absence of d-Gal, and BH→SA-Cy3 labeling. (e) CLSM images of spermatozoa with or without pretreatment by neuraminidase (NEU) followed by NaIO4 oxidation and BH→SA-Cy3 labeling. Cell nuclei were stained with DAPI (blue). Scale bars: 5 µm. The above data are representative of three individual experiments.
Figure 2. (a) Schematic illustration of the tandem glycan chemoselective labeling technique. (b) CLSM images of spermatozoa after different treatment combinations shown in (a). Cell nuclei were stained with DAPI (blue). Scale bar: 5 µm. All images are representative of three individual experiments.
Figure 3. (a) Schematic showing the labeling protocol of spermatozoa from clinical normal and abnormal human semen samples. (b) Representative CLSM images and (c) flow cytometric results of normal and abnormal human spermatozoa undergoing CGL and fluorescence staining. Scale bar: 5 µm. (d) Quantification of median fluorescence intensity (MFI) in (c). The expression level of glycans (Gal/GalNAc or Sia) in abnormal spermatozoa relative to that of normal spermatozoa was shown. Data are shown as the mean ± SEM, n = 10 (t-test). P < 0.05; **P < 0.01.
Figure 4. (a) Schematic showing the labeling protocol of spermatozoa from normal (BKS) and diabetic (db/db) mice. (b) Representative CLSM images and (c) flow cytometric results of BKS and db/db mice spermatozoa undergoing CGL labeling and fluorescence staining. Scale bar: 5 µm. (d) Quantification of MFI in (c), n = 13 (t-test). The expression level of glycans (Gal/GalNAc or Sia) in spermatozoa from diabetic mice relative to that from normal mice was shown. (e) Spermatozoa density of BKS mice and db/db mice (n = 5 (t-test)) are shown as points in the boxplots, the center lines represent the median, box edges show the 25th and 75th percentiles, and whiskers represent min to max. (f) Motility detection of spermatozoa from BKS mice and db/db mice, n = 5 (t-test). Data in (d) and (f) are shown as the mean ± SEM. P < 0.05; **P < 0.01; ***P < 0.001.

 Login In
Login In




 DownLoad:
DownLoad:
 DownLoad:
DownLoad: